
公司动态首页 > 新闻动态 > 公司动态
背景介绍
结核病(Tuberculosis)俗称“痨病”,是由结核分枝杆菌(Mycobacterium tuberculosis,Mtb)侵入人体后引起的一种慢性传染病,其本质是结核分枝杆菌与人体免疫系统相互斗争的结果,全身各脏器(除头发、指甲外)都可发病,以肺部为主。
吡嗪酰胺(PZA)是治疗结核病的关键药物,在结核分枝杆菌体内通过吡嗪酰胺酶转化成活性成分吡嗪酸,可实现结核病的有效治疗。但是,吡嗪酸在结核分枝杆菌中会出现外排现象,从而影响药效。复旦大学华山医院张颖教授团队和张文宏教授团队合作[1],对此做了深入的研究。他们利用博翀生物的结核分枝杆菌蛋白(MtbProt)芯片, 找到Mtb的吡嗪酸外排结合蛋白Rv0191、Rv3756c、Rv3008和Rv1667c。
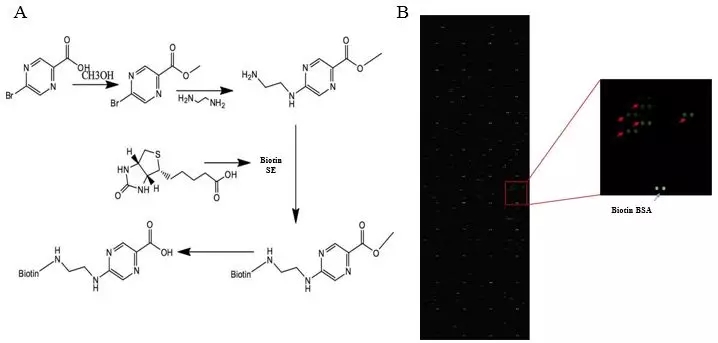
图1. 吡嗪酸生物素标记及芯片结果展示图
Mtb的耐药机制极为复杂,主要包括结核菌细胞壁结构与组成变化[2-3]、自身药物外排泵系统[5]、药物靶编码基因发生突变[6-7]等。针对不同种类抗生素,Mtb有不同的耐药机制,而对同一种抗生素也可能存在多种耐药机制[4]。随着临床新型抗菌药物的研发和使用,新的耐药机制也在不断被发现。及时掌握Mtb的耐药情况变化,对合理使用抗菌药物和更好地控制Mtb耐药性的发生具有重要意义。
小分子药物对结核病的治疗是一个重要的开发领域,不管是已有治疗结核病药物的耐药机制,还是新发现的活性小分子的作用机制,最后都要聚焦于药物靶标蛋白上。利用结核杆菌蛋白芯片可以快速找到小分子直接作用靶标,指导后续的功能研究以及提供潜在的药物靶标。
MtbProt芯片的药物靶蛋白筛选方案
1. 结核分枝杆菌蛋白芯片简介
全球第一张结核杆菌全蛋白芯片由中国科学院生物物理所和上海交通大学合作构建,这也是全球目前唯一一种基于重组蛋白的结核杆菌蛋白芯片产品。该结核杆菌蛋白芯片(MTB proteome microarray TM)涵盖了95%的结核杆菌,共4262个蛋白质,并且在项目合作单位广州博翀生物科技有限公司实现商业化。该芯片在寻找蛋白靶标[8-9]、小分子靶标[10-12]等方面发表了一系列文章,结合人类蛋白质组芯片[13],可以研究病原宿主相互作用网络。
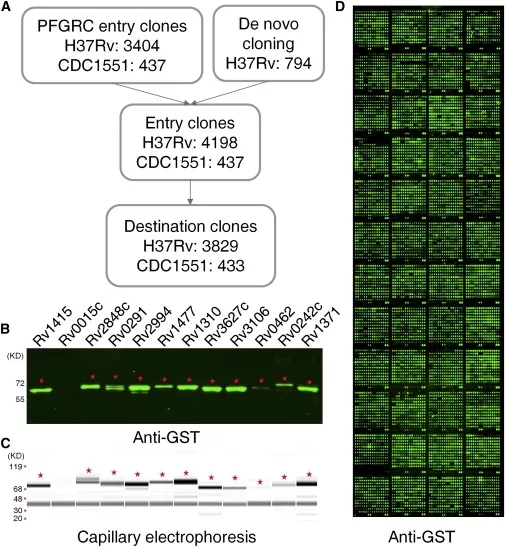
图2. 结核杆菌蛋白芯片的构建及质控图
2. 实验设计
(1)小分子进行生物素标记(含有游离的羟基、羧基、氨基;或者多步反应);
(2)生物素标记好的小分子进行芯片前的活性验证(和未标记小分子比较);
(3)标记好的小分子与结核杆菌蛋白芯片孵育、清洗后,芯片扫描仪解读芯片数据;
(4)设置合适cutoff,得到潜在蛋白并数据处理,GO分析、pathway分析;
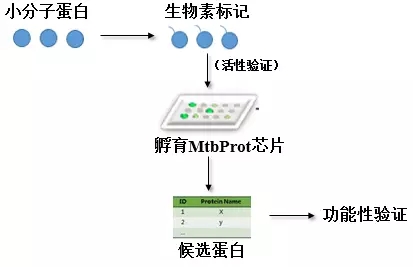
图3. 实验设计流程图
拓展延伸
博翀生物的合作伙伴拥有成熟的结核分枝杆菌基因敲除技术,截至目前已经构建噬菌体文库及200多个结核分枝杆菌基因敲除菌株,可为客户提供结核分枝杆菌基因敲除相关产品及技术服务。此外,他们还构建了结核分枝杆菌基因过表达质粒及过表达菌株文库,拥有荧光标记菌株构建技术。
欢迎联系我们,博翀生物携手晶诺生物为客户科研提供服务,助力结核病基础研究。
博翀生物MtbProt®结核分枝杆菌蛋白芯片:

参考文献
[1] Zhang Y, et al. Identification of novel efflux proteins Rv0191, Rv3756c, Rv3008 and Rv1667c involved in pyrazinamide resistance in Mycobacterium tuberculosis [J]. Antimicrob Agents Chemother, 2017, 61(8):AAC.00940-17.
[2] Alderwick L J, et al. Identification of a Novel Arabinofuranosyltransferase (AftA) Involved in Cell Wall Arabinan Biosynthesis in Mycobacterium tuberculosis [J]. Journal of Biological Chemistry, 2006, 281(23):15653-15661.
[3] Seidel M, et al. Identification of a novel arabinofuranosyltransferase AftB involved in a terminal step of cell wall arabinan biosynthesis in Corynebacterianeae, such as Corynebacterium glutamicum and Mycobacterium tuberculosis [J]. Journal of Biological Chemistry, 2007, 282(20):14729-14740.
[4] Suárezgarcía I, et al. Risk factors for multidrug-resistant tuberculosis in a tuberculosis unit in Madrid, Spain [J]. Eur J Clin Microbiol Infect Dis, 2009, 28(4):325-330.
[5] Li X Z, et al. Efflux Pump-Mediated Intrinsic Drug Resistance in Mycobacterium smegmatis [J]. Antimicrobial Agents & Chemotherapy, 2004, 48(7):2415-23.
[6] Mukherjee A, et al. Changing trends in childhood tuberculosis [J]. Indian Journal of Pediatrics, 2011, 78(3):328-333.
[7] Yin X, Yu Z. Mutation characterization of gyrA and gyrB genes in levofloxacin-resistant Mycobacterium tuberculosis clinical isolates from Guangdong Province in China [J]. Journal of Infection, 2010, 61(2):150-154.
[8] Deng J, et al. Mycobacterium tuberculosis proteome microarray for global studies of protein function and immunogenicity [J]. Cell Reports, 2014, 9(6):2317-2329.
[9] Wu F L, et al. The Ser/Thr protein kinase protein-protein interaction map of M. tuberculosis [J]. Molecular & Cellular Proteomics Mcp, 2017, 16(8):mcp.M116.065771.
[10] He X, et al. Systematic identification of Mycobacterium tuberculosis effectors reveals that BfrB suppresses innate immunity [J]. Molecular & Cellular Proteomics, 2017, 16(12):mcp.RA117.000296.
[11] Zhang H N, et al. Cyclic di-GMP regulates Mycobacterium tuberculosis resistance to ethionamide [J]. Scientific Reports, 2017, 7(1):5860.
[12] Cheng X, et al. Proteomic identification of the oncoprotein STAT3 as a target of a novel Skp1 inhibitor [J]. Oncotarget, 2016, 8(2):2681.
[13] Jing W, et al. The ubiquitin ligase TRIM27 functions as a host restriction factor antagonized by Mycobacterium tuberculosis PtpA during mycobacterial infection [J]. Scientific Reports, 2016, 6:34827.
博翀生物期待和您在抗结核药物靶蛋白筛选方面进行深度合作,欢迎来电来函咨询。
联系电话:4008346886;客服邮箱:info@bcbio.cn。

